
Energy = The Primary Currency of Life
The efficiency with which an individual acquires, converts, and expends energy greatly influences its fitness. I integrate a variety of field technologies with modeling techniques to understand how species pay the various costs associated with life. Key topics of interest include:
Efficient Movement
Locomotion is a fundamental biological process, especially for animals that must travel vast distances in search of food, mates, or suitable habitats. I investigate how body size, morphology, and biomechanics influence movement efficiency across a range of species. By integrating high-resolution motion data with kinematic models, I explore how animals generate propulsion, maneuver through their environment, and optimize their energy use.
Finding and Capturing Food
Energy acquisition is fundamental to survival, and animals have evolved diverse strategies to locate, capture, and process food while minimizing costs. I study how species optimize their foraging efficiency through behavioral adaptations, biomechanical performance, and environmental interactions. By integrating drone-based videography, biologging tags, and computational models, I analyze how movement patterns, prey capture techniques, and metabolic demands shape energy intake at both individual and population levels.

Figure from Gough et al. 2025 (in review at JEB) - Overview of a single foraging dive performed by a short-finned pilot whale tagged with a CATS biologging tag. Part (A) shows the depth (top), swimming speed (middle), and per-second energetic expenditure (bottom) for the full period of the dive. Blue segments along the depth trace represent acoustic click production, while orange circles denote the timing of foraging buzzes derived from acoustic data. Gray segments correspond with periods (>10 s) of active swimming; white corresponds with un-powered gliding. Black circles along the top denote the start times of individual tailbeats. Fine-scale energetic expenditure estimates from the thrust method (solid line) are shown in comparison to the simplified breathing frequency method (dotted line). Part (B) shows the acoustic record (top), jerk (middle), and swimming speed (bottom) for a zoomed-in segment of data around three foraging buzzes derived from acoustic data. Orange dotted lines denote the end positions of each buzz. Part (C) shows a further zoomed-in period of the acoustic record around one of the foraging buzzes (Buzz 2). Parts (D-F) show images from the CATS video record before, during, and immediately after the dive. The images in (E) correspond to Buzz 2, showing the ocean floor and what appears to be a squid ink cloud appearing <2 s from the end time of the buzz.
Balancing Energy Budgets
Survival depends on maintaining a delicate balance between energy intake and expenditure. Whether foraging, migrating, or reproducing, animals must allocate energy efficiently to meet their physiological demands. I examine how metabolic constraints, movement patterns, and environmental variability influence energy budgeting across different life stages and ecological contexts. By combining empirical data with bioenergetic models, I aim to understand how animals manage trade-offs between energy acquisition and expenditure.
Scaling and Ecological Consequences
Body size fundamentally shapes how animals move, feed, and interact with their environment. I explore how biomechanical performance and energetic efficiency scale across species, revealing broader patterns in ecological specialization and adaptation. By investigating these relationships across diverse taxa, I aim to uncover fundamental principles that govern how energy use and movement strategies evolve in response to ecological pressures.
Current Ongoing Projects
CATS tag image from bubble-netting whale in southeastern Alaska. The tag is deployed on the dorsum and oriented forward, with the left pectoral flipper raised and a stream of bubbles appearing out of the blowhole.
Kinematics and Energetics of Bubble-Net Feeding In Southeast Alaskan Humpbacks
Bubble-netting is a highly specialized foraging behavior employed by humpback whales (Megaptera novaeangliae) in multiple locations around the globe. All forms of bubble-netting involve the use of a bubble curtain to corral or startle prey into a dense aggregation for easier consumption at higher densities, thereby increasing the energetic economy of the foraging bout. Different populations of animals have developed these strategies independently over time and use their bubble curtain in different ways. Some groups forage solitarily or in pairs and create a series of successively smaller curtains before lunging, while other groups range upwards in size from ~4-16 individuals, with designated “bubble-blowers” that form the bubble curtain. Many of these bubble-netting variations have been studied individually, but our study will be the first to directly compare the kinematics and energetic economy of different bubble-netting styles using a combination of UAV drone and biologging tag data for animals from three distinct populations.
Representative annual cycle for a mysticete whale (Humpback: Megaptera Novaeangliae). The top left depicts the variables affecting daily foraging variability. The bottom left depicts a migration between a foraging ground (Western Antarctic Peninsula) and a breeding ground (Ecuador). The right side depicts the annual cycle of energy loss and energy gain.
Migration Energetics of large Whales
Long-distance migrations are one of the most energetically challenging behaviors observed within the animal kingdom. Animals migrate to track the seasonality of food sources or move between spatially distinct foraging and breeding habitats. Large body sizes, such as those seen in baleen whales (Mysticeti), are one adaptation allowing for migrations across vast distances. As capital breeders, mysticetes must also rely on the energy from a defined feeding season to last them throughout the year. Consequently, maximum migratory distance should be a function of energy gained during the feeding season. Using empirically-derived estimates of foraging and swimming energetic intake and costs for mysticetes of varying body sizes, we calculated the energy gained during a foraging season as well as the energy subsequently used during migration. For a successful foraging season, the energetic cost of migration only amounts to ~20% of foraging season energy intake across body sizes, while a poor foraging year could result in exorbitant migratory costs of ~100% or more. We also found that migratory costs are dependent on total distance, duration, and swimming speed. Combining a theoretical model with satellite tracks of individual whales, we determined that longer migrations are more costly and occur at higher speeds than shorter migrations. In a rapidly changing ocean, small differences in the distance, duration, and/or speed of a migration could have major impacts on individual fitness.
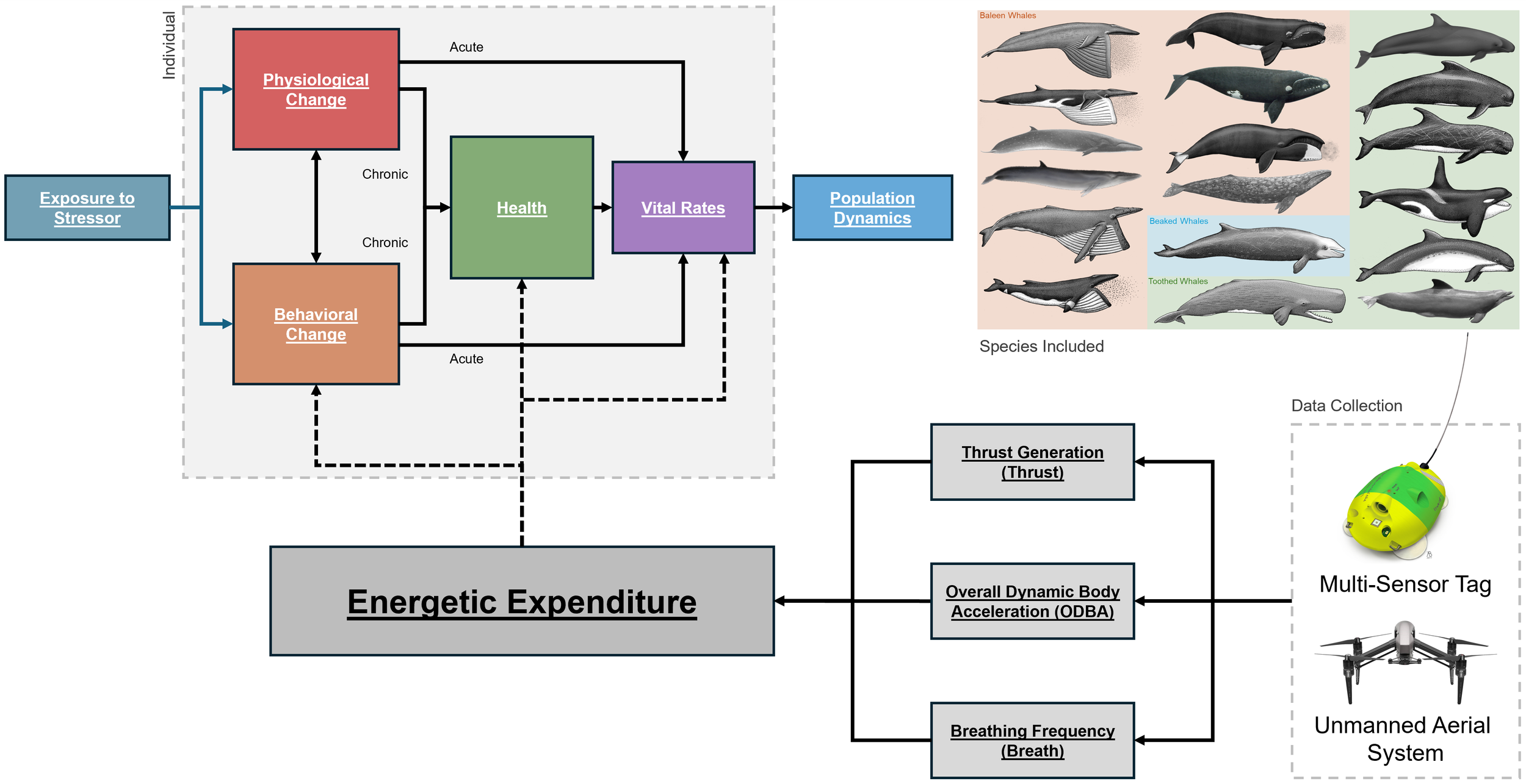
Population Consequences of Disturbance (PCOD) model (modified from Pirotta et al. 2018). Estimates of energetic expenditure are expected to inform the “Behavioral Change”, “Health”, and “Vital Rates” portions of the PCOD model. Data collection avenues (Multi-Sensor Tag and Unmanned Aerial System) and methods of calculating energetic expenditure (Thrust Generation, Overall Dynamic Body Acceleration, Breathing Frequency) are outlined. Illustrations show the 18 species to be included in methods comparisons - pink are baleen whales, blue is a beaked whale, and green are other toothed whales.
Energetic Expenditure of Cetaceans Across Scale
Energy use underpins every aspect of cetacean ecology, from foraging and migration to reproduction and survival. In this project, we are synthesizing large, existing datasets to better understand how energetic expenditure varies across whale and dolphin species and how it is best quantified in free-ranging animals. We are integrating on-animal biologging and drone-based photogrammetry data from hundreds of tag deployments spanning 18 cetacean species, representing a broad range of body sizes, life histories, and ecological settings. By comparing multiple approaches used to estimate energetic expenditure, we evaluate how methodological choices influence energetic estimates and how those estimates scale across species and behaviors. This work provides foundational energetic benchmarks needed to link short-term behavioral responses to long-term population outcomes. The results directly support Population Consequences of Disturbance (PCOD) frameworks and improve our ability to assess how environmental change and human activities influence marine mammal health, resilience, and population dynamics.

2) Cade, D.E., Kahane-Rapport, S.R., Gough, W.T., Bierlich, K.C., Linsky, J.M.J., Calambokidis, J., Johnston, D.W., Goldbogen, J.A. and Friedlaender, A.S. (2023). Minke whale feeding rate limitations suggest constraints on the minimum body size for engulfment filtration feeding. Nature Ecology & Evolution 7: 535-546.
3) Segre, P.S., di Clemente, J., Kahane-Rapport, S.R., Gough, W.T., Meÿer, M.A., Lombard, A.T., Goldbogen, J.A. and Penry, G.S. (2022). High-speed chases along the seafloor put Bryde’s whales at risk of entanglement. Conservation Science and Practice 4: e12646.
4) Gough, W.T., Cade, D.E., Czapanskiy, M.F., Potvin, J., Fish, F.E., Kahane-Rapport, S.R., Savoca, M.S., Bierlich, KC, Johnston, D.W., Friedlaender, A.S., Szabo, A., Bejder, L. and Goldbogen, J.A. (2022). Fast and furious: energetic tradeoffs and scaling of high-speed foraging in rorqual whales. Integrative and Organismal Biology 4: obac038.
5) Segre, P.S., Gough, W.T., Roualdes, E.A., Cade, D.E., Czapanskiy, M.F., Fahlbusch, J., Kahane-Rapport, S.R., Oestreich, W.K., Bejder, L., Bierlich, K.C., Burrows, J.A., Calambokidis, J., Chenoweth, E.M., Di Clemente, J., Durban, J.W., Fearnbach, H., Fish, F.E., Friedlaender, A.S., Hegelund, P., Johnston, D.W., Nowacek, D.P., Oudejans, M.G., Penry, G.S., Potvin, J., Simon, M., Stanworth, A., Straley, J.M., Szabo, A., Videsen, S.K.A., Visser, F., Weir, C.R., Wiley, D.N. and Goldbogen, J.A. (2022). Scaling of maneuvering performance in baleen whales: larger whales outperform expectations. Journal of Experimental Biology 225: jeb243224.
6) Cade, D.E., Gough, W.T., Czapanskiy, M.F., Fahlbusch, J.A., Kahane-Rapport, S.R., Linsky, J.M.J., Nichols, R.C., Oestreich, W.K., Wisniewska, D.M., Friedlaender, A.S. and Goldbogen, J.A. (2021). Tools for integrating inertial sensor data with video bio-loggers, including estimation of animal orientation, motion, and position. Animal Biotelemetry 9: 1-21.

7) Savoca, M.S., Czapanskiy, M.F., Kahane-Rapport, S.R., Gough, W.T., Fahlbusch, J.A., Bierlich, KC, Segre, P.S., Di Clemente, J., Penry, G.S., Wiley, D.N., Calambokidis, J., Nowacek, D.P., Johnston, D.W., Pyenson, N.D., Friedlaender, A.S., Hazen, E.L. and Goldbogen, J.A. (2021). Baleen whale prey consumption based on high resolution foraging measurements. Nature 599: 85-90.
8) Modest, M., Irvine, L., Andrews-Goff, V., Gough, W.T., Johnston, D., Nowacek, D., Pallin, L., Read, A., Moore, R.T. and Friedlaender, A. (2021). First description of migratory behavior of humpback whales from an Antarctic feeding ground to a tropical calving ground. Animal Biotelemetry 9: 42.
9) Czapanskiy, M.F., Savoca, M.S., Gough, W.T., Segre, P.S., Wisniewska, D.M., Cade, D.E. and Goldbogen, J.A. (2021). Modelling short-term energetic costs of sonar disturbance to cetaceans using high-resolution foraging data. Journal of Applied Ecology 58: 1643-1657.
10) Fish, F.E., Sheehan, M.J., Adams, D.S., Tennett, K.A. and Gough, W.T. (2021). A 60:40 split: differential mass support in dogs. Anatomical Record 304: 78-89.
11) Gough, W.T., Smith, H.J., Savoca, M.S., Czapanskiy, M.F., Fish, F.E., Potvin, J., Bierlich, KC, Cade, D.E., Di Clemente, J., Kennedy, J., Segre, P.S., Stanworth, A., Weir, C.R. and Goldbogen, J.A. (2021). Scaling of oscillatory kinematics and Froude efficiency in baleen whales. Journal of Experimental Biology 224: jeb237586.
12) Segre, P.S., Potvin, J., Cade, D.E., Calambokidis, J., Di Clemente, J., Fish, F.E., Friedlaender, A.S., Gough, W.T., Kahane-Rapport, S.R., Oliveira, C., Parks, S.E., Penry, G.S., Simon, M., Stimpert, A.K., Wiley, D.N., Bierlick, KC, Madsen, P.T. and Goldbogen, J.A. (2020). Energetic and physical limitations on the breaching performance of large whales. eLife 9.

13) Goldbogen, J.A., Cade, D.E., Wisniewska, D.M., Potvin, J., Segre, P.S., Savoca, M.S., Hazen, E.L., Czapanskiy, M.F., Kahane-Rapport, S.R., DeRuiter, S.L., Gero, S., Tønnesen, P., Gough, W.T., Hanson, M.B., Holt, M.M., Jensen, F.H., Simon, M., Stimpert, A.K., Arranz, P., Johnston, D.W., Nowacek, D.P., Parks, S.E., Visser, F., Friedlaender, A.S., Tyack, P.L., Madsen, P.T. and Pyenson, N.D. (2019). Why whales are big but not bigger: Physiological drivers and ecological limits in the age of ocean giants. Science 366: 1367–1372.
14) Goldbogen, J.A., Cade, D.E., Calambokidis, J., Czapanskiy, M.F., Fahlbusch, J.A., Friedlaender, A.S., Gough, W.T., Kahane-Rapport, S.R., Savoca, M.S., Ponganis, K.V. and Ponganis, P.J. (2019). Extreme bradycardia and tachycardia in the world’s largest animal. Proceedings of the National Academy of Sciences 4: 25329-25332.
15) Gough, W.T., Segre, P.S., Bierlich, KC, Cade, D.E., Potvin, J., Fish, F.E., Dale, J., Di Clemente, J., Friedlaender, A.S., Johnston, D.W., Kahane-Rapport, S.R., Kennedy, J., Long, J.H., Oudejans, M., Penry, G., Savoca, M.S., Simon, M., Videsen, S.KA., Visser, F., Wiley, D.N. and Goldbogen, J.A. (2019). Scaling of swimming performance in baleen whales. Journal of Experimental Biology 222: jeb204172.
16) Gough, W.T., Fish, F.E., Wainwright, D.K. and Bart-Smith, H. (2018). Morphology of the core fibrous layer of the cetacean tail fluke. Journal of Morphology 279: 757-765.
17) McGuire, B. and Gough, W.T. (2017). Body size influences urinary posture but not hindlimb laterality in shelter dogs. Journal of Veterinary Behavior: Clinical Applications and Research 21: 38-44.
18) Gough, W.T. and McGuire, B. (2015). Urinary Posture and Motor Laterality in Dogs (Canis lupus familiaris) at Two Shelters. Applied Animal Behaviour Science 168: 61-70.
19) Gough, W.T., Farina, S.C. and Fish, F.E. (2015). Aquatic Burst Locomotion by Hydroplaning and Paddling in Common Eiders (Somateria mollissima). Journal of Experimental Biology 218: 1632-1638.
TEACHING
Marine Mammal Science and Conservation (MBIO 650) (~6 students) – 2023
• University of Hawaii at Manoa (HIMB) led by Dr. Lars Bejder
• Led two days of teaching / training related to the history and modern usage of biologging technologies.
Biologging Tag Data Processing Workshop (~50 attendees) – (2020)
• Stanford University / UC Santa Cruz in collaboration with Dr. David Cade.
• Co-designed / co-instructed a week-long workshop to train members of the scientific community to 1) properly setup and deploy archival biologging tags, 2) process the resulting data into useable formats, and 3) design experiments to answer novel biomechanical, behavioral, or physiological questions.
Biologging and Biotelemetry (BIOHOPK 234H) (~10 students) – (2018)
• Stanford University (Hopkins Marine Station) led by Dr. Jeremy Goldbogen.
• Assisted with class preparation and activities to train students on the proper use and potential utility of biologging devices.
Animal and Plant Physiology (BIO 84) (~40 students) – (2018)
• Stanford University led by Drs. Craig Heller, Robert Sapolsky, and Jose Dinneny.
• Led two weekly discussion sessions to reinforce lecture material focused on multiple physiological processes in plant and animal systems.
• Proctored exams and assisted with grading of all class materials.
Anatomy and Function of Marine Vertebrates (BIOSM 3210) – (2013 – 2014)
• Cornell University (Shoals Marine Laboratory) led by Dr. Frank Fish
• Helped students develop and complete individual, research-based final projects pertaining to multiple marine vertebrate lineages.
• Prepared lab and dissection materials and assisted with grading class materials.
Vertebrate Anatomy, Physiology, and Evolution (BIOEE 2740) – (2013)
• Cornell University led by Dr. Betty McGuire.
• Assisted with lab material preparation and proctoring of laboratory examinations.
OUTREACH
Stanford University – Salinas High School Internship Program – (2019 – 2022)
• Co-led design and logistics for a partnered internship between Hopkins Marine Station and Salinas High School.
• Managed pairings of 8-10 high school interns per year with graduate student / post-doctoral mentors.
• Designed final seminars to give interns a chance to present their work to their peers and parents.
Pacific Grove Natural History Museum – Science Saturday – (2019)
• Presented cetacean research to a public audience of adults and children
American Museum of Natural History – Milstein Science Series – (2015)
• Presented simple marine biomechanics and locomotion concepts to a public audience of adults and children.
Cornell University – Expand Your Horizons – (2013 – 2014)
• Assisted and led scientific activities for a group of 15-20 7th grade girls.